
a special guest post by John Guinotte, Marine Conservation Biology Institute

The answer is uncertain as very few manipulative experiments have been conducted to test how deep-sea corals react to changes in temperature, seawater chemistry (pH), water motion (currents), and food availability. It is likely that the effects of climate change will not be positive for deep-sea corals, because they are highly specialized and have evolved under very stable (cold, dark, nutrient-rich) conditions. Temperature, salinity, seawater chemistry, and light availability control calcification rates in shallow, tropical reef systems and with the exception of light (because deep-sea corals lack algal symbionts), these factors are probably important controls on deep-sea coral calcification as well.
Seawater Chemistry (pH)
Seawater chemistry and the calcium carbonate saturation state of the world’s oceans are changing with the addition of fossil fuel CO2 to the atmosphere (Feely et al., 2004). This influx of CO2 is causing the world’s oceans to become more acidic, which is bad news for corals and other marine organisms that use calcium carbonate to build their skeletons and protective shells. Although little is known about the effects of decreasing carbonate saturation state on deep-sea corals, lab experiments have conclusively shown that lowering carbonate ion concentration reduces calcification rates in tropical reef builders by 7 to 40 percent (Kleypas et al 2006; Marubini et al., 2003; Langdon et al., 2000; Gattuso et al., 1999). In fact, all marine calcifying organisms tested to date have shown a similar negative response. As the world’s oceans become less saturated over time, corals are expected to build weaker skeletons (a process similar to osteoporosis in humans) and/or to experience slower growth rates. If saturation state is as important to deep-sea coral calcification as it is to shallow reef builders, then this is an important issue. Currently, greater than 95% of structure forming deep sea scleractians are found in waters that are supersaturated with respect to aragonite. By 2100, 70% of these coral locations will be in undersaturated water (Guinotte et al 2006). Cairns (2007) points out that the depth of the calcium carbonate saturation horizons could be limiting deep-sea coral diversity in some regions of the world’s oceans.
Temperature and Dissolved Oxygen
Global sea temperatures are rising both in surface waters and in the deep sea due to an influx of anthropogenic carbon dioxide to the atmosphere. Since the industrial revolution, there has been a documented increase in shallow water coral bleaching events, and in many cases, massive coral mortality has followed. Deep-sea corals cannot bleach, since they lack symbiotic algae, but rising temperatures will probably influence their calcification rates, physiology, and biochemistry. Deep-sea corals grow very slowly (4 to 25 mm/yr or less; Freiwald et al., 2004), an order of magnitude slower than tropical corals. This difference is probably due to the cold temperatures in which deep-sea corals live (about 4° to 13°C). Preliminary findings suggest Lophelia pertusa
respiration rates increase when the coral is exposed to higher ambient temperatures, but the synergistic effects of increased respiration, temperature, and PCO2 on calcification are not known (Dodds et al 2007). Dodds et al (2007) also identify dissolved oxygen concentration as a possible limiting factor in the distribution of the structure forming coral L. pertusa. This could potentially be an emerging issue as climate models predict an overall decline in oceanic dissolved oxygen concentration under global warming conditions (Stramma et al 2008, Matear and Hirst 2003)
Food Availability
Very little information exists on the food sources of deep-sea corals. It is probable that they depend on suspended organic matter and zooplankton for nourishment (Freiwald et al., 2004). Since corals are sessile filter-feeding organisms, they can obtain nourishment either from organic matter falling from the surface or via currents that bring organic matter and zooplankton to the coral. Deep-sea corals occur in waters that have higher than average surface primary productivity, indicating that food falling from the surface is important to their survival (Guinotte et al 2006). Many species of plankton (e.g., coccolithophores and foramniferans) build calcium carbonate shells and are sensitive to the seawater chemistry changes previously noted. If ecosystem changes due to climate change reduce surface productivity, food sources for deep-sea corals will probably also be reduced.
Summary
Deep-sea coral ecosystems have probably not experienced a combination of stresses of the types described above for a very long time. The synergistic effects of these stresses occurring in concert are uncertain, but changes in the environmental factors identified above will probably have serious implications for deep-sea coral ecosystems. In situ monitoring and lab experiments are needed to help us understand and quantify how these synergistic changes might affect deep-sea coral ecosystems.
Modified from:
Guinotte JM (2005) Climate change and deep-sea corals. Current: The Journal of Marine Education 21(4):48-49.
References:
Cairns, S. 2007. Deep-water corals: an overview with special reference to diversity and distribution of deep-water scleractinian corals. Bulletin of Marine Science, 81(3): 311-322.
Dodds LA, Roberts JM, Taylor AC, Marubini F (2007) Metabolic tolerance of the cold-water coral Lophelia pertusa (Scleractinia) to temperature and dissolved oxygen change. Journal of Experimental Marine Biology and Ecology 349: 205-214
Feely, R., C. Sabine, K. Lee, W. Berelson, J. Kleypas, V. Fabry,and F. Millero (2004). “Impact of Anthropogenic CO2 on the CaCO3 System in the Oceans.” Science 305:362-366.
Freiwald, A., J.H. Fosså, A. Grehan, T. Koslow, and J.M. Roberts (2004). Cold-Water Coral Reefs. UNEP-WCMC, Cambridge,UK.
Gattuso, J., D. Allemand, and M. Frankignoulle (1999). “Photosynthesis and Calcification at Cellular, Organismal and Community Levels in Coral Reefs: A Review on Interactions
and Control by Carbonate Chemistry,” American Zoologist 39:160-183.
Guinotte, J. M. et al. 2006. Will human induced changes in seawater chemistry alter the
distribution of deep-sea scleractinian corals? Front Ecol Environ 4: 141-146.
Kleypas, J. A., et al. 2006. Impacts of Ocean Acidification on Coral Reefs and Other
Marine Calcifiers: A Guide for Future Research, report of a workshop held 18-20
April 2005, St. Petersburg, FL, sponsored by NSF, NOAA, and the U.S. Geological Survey, 88 pp.
Langdon, C., T. Takahashi, C. Sweeney, D. Chipman, and J. Goddard (2000). “Effect of Calcium Carbonate Saturation State on the Calcification Rate of An Experimental Coral Reef.” Global Biogeochemical Cycle 14(2):639-654.
Marubini, F., C. Ferrier-Pages, J.P. Cuif (2003). “Suppression of Skeletal Growth in Scleractinian Corals by Decreasing Ambient Carbonate-Ion Concentration: A Cross-Family Comparison.” Proceedings of the Royal Society of London, Series B 270: 179-184.
Matear, R.J., and A. C. Hirst. (2003) Global Biogeochem. Cycles 17, 10.1029/2002GB001997.
Stramma et al. 2008. Expanding oxygen-minimum zones in the tropical oceans. Science 320: 655-658.

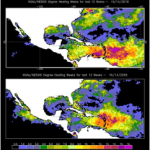


Jim, what cites can one show the people who dispute reports that “Global sea temperatures are rising both in surface waters and in the deep sea” — I have seen a very few papers and more abstracts (as a non-academic, I can’t read most journal articles behind paywalls) but don’t know which ones are cited as reliable in the field. I realize measurements are thin yet although the physics says what to expect.